Nature背靠背 | 终于搞清楚了!破解METTL1-WDR4复合物如何催化tRNA m7G修饰
时间:2023-01-08 15:00:48 热度:37.1℃ 作者:网络
RNA的化学修饰在许多生物过程中起着关键作用。tRNA富含各种化学修饰,这些修饰影响其稳定性和功能,其中N7-甲基鸟苷(N7-methylguanosine, m7G)是tRNA的完整性和稳定性所必需的。
tRNA m7G甲基化复合物主要有甲基化酶METTL1和辅助蛋白WDR4组成。它在某些tRNAs的可变环中修饰G46,其调控异常在许多癌症类型中驱动肿瘤发生。WDR4突变导致包括小头症在内的人类发育表型。然而,METTL1-WDR4是如何修饰tRNA底物并被调节的仍然是难以捉摸的。
2023年1月4日,美国波士顿儿童医院Richard I. Gregory课题组在Nature杂志在线发表题为“Structural basis of regulated m7G tRNA modification by METTL1-WDR4”的研究论文,该研究通过对人类METTL1-WDR4的结构、生化和细胞研究表明,WDR4可作为METTL1和tRNA T臂的支架。在tRNA结合后,METTL1的αC区转变为一个螺旋,该螺旋与α6螺旋一起固定tRNA变量环的两端。研究发现METTL1预测的无序N端区域是催化口袋的一部分,对甲基转移酶活性至关重要。此外,研究发现S27在METTL1 N端区域的磷酸化通过局部破坏催化中心来抑制甲基转移酶活性。总之,这项研究结果提供了tRNA底物识别和磷酸化介导的METTL1-WDR4调控的分子理解,并揭示了METTL1无序的N端区域作为甲基转移酶活性的连接。
另外,2023年1月4日,德克萨斯大学西南医学中心Yunsun Nam课题组背靠背在Nature 发表题为“Structures and mechanisms of tRNA methylation by METTL1–WDR4”的研究论文,该研究展示了METTL1和WDR4如何合作识别RNA底物和催化甲基化。METTL1-WDR4的晶体结构和冷冻电镜结构表明,复合蛋白表面通过形状互补识别tRNA弯头。METTL1-WDR4-tRNA与S-腺苷甲硫氨酸或S -腺苷同型半胱氨酸的冷冻电镜结构以及METTL1晶体结构通过揭示多种状态下的活性位点,为催化机制提供了额外的见解。METTL1 N端与tRNA、催化环和WDR4 C端构象变化的辅因子结合,作为激活m7G甲基化的开关。因此,这些结构模型解释了METTL1 N端翻译后修饰如何调节甲基化。总之,这项工作阐明了METTL1修饰m7G的核心和调控机制,为理解其对生物学和疾病的贡献提供了框架。


最近的“外延转录组”研究揭示了RNA修饰在各种分子、细胞和发育过程中的关键作用。一些儿童发育障碍是由编码RNA修饰酶的基因突变引起的,某些RNA修饰的失调是致癌的。
tRNA是修饰程度最高的一类RNA,影响tRNA的成熟、稳定性和功能。其中,位于G46位置的m7G (m7G46)是大量tRNAs中普遍存在的修饰。m7G46在D-loop中与C13-G22相互作用以稳定tRNA三级结构。缺少m7G46与tRNA快速衰减和表达改变相关。m7G46由METTL1及其必要辅因子WDR4催化。METTL1-WDR4复合物是正常mRNA翻译和小鼠胚胎干细胞增殖和分化所必需的。WDR4突变与发育缺陷相关,包括原发侏儒症、小头畸形症和 Galloway–Mowat综合征。失调的METTL1-WDR4驱动肿瘤发生,这种致癌功能与许多不同的癌症类型有关。然而,缺乏人类METTL1-WDR4的结构和机制信息仍然是阐明m7G46修饰过程和开发甲基转移酶(MTase)抑制剂作为可能的新型抗癌药物的主要障碍。
m7G46修饰发生在变量loop中包含' RAGGU '基序的大量tRNAs中。体外MTase测试显示了酵母Trm8-Trm82复合物(分别是METTL1和WDR4的同源物)对m7G修饰的D臂和T臂的重要性。tRNA识别模型之前是通过计算将tRNA结构与Trm8-Trm82的结构对接而生成的。该模型表明tRNA与Trm8的正电荷表面相互作用而不与Trm82接触,并且仅基于酵母复合物的部分结构,该结构缺乏m7G催化所需的区域。因此,m7G MTase复合物如何特异性地接合和修饰tRNA底物仍然未知。
通过蛋白激酶B(protein kinase B, 又名AKT)磷酸化METTL1 (S27)可以通过一种未知的机制使MTase活性失活。值得注意的是,S27位于METTL1的N端,其结构信息是不可用的,被预测为很大程度的无序。然而,灵活的METTL1 N端是保守的,这意味着重要的功能。
该研究报道了人类METTL1-WDR4异源二聚体的结构,并与tRNA底物tRNAValTAC和tRNAPheGAA配合物。不同的tRNA表现出与METTL1-WDR4相似的结合模式,其中MTase复合物和tRNA的结构重排都是可见的。结合功能分析揭示了METTL1-WDR4修饰m7G46的分子机制。
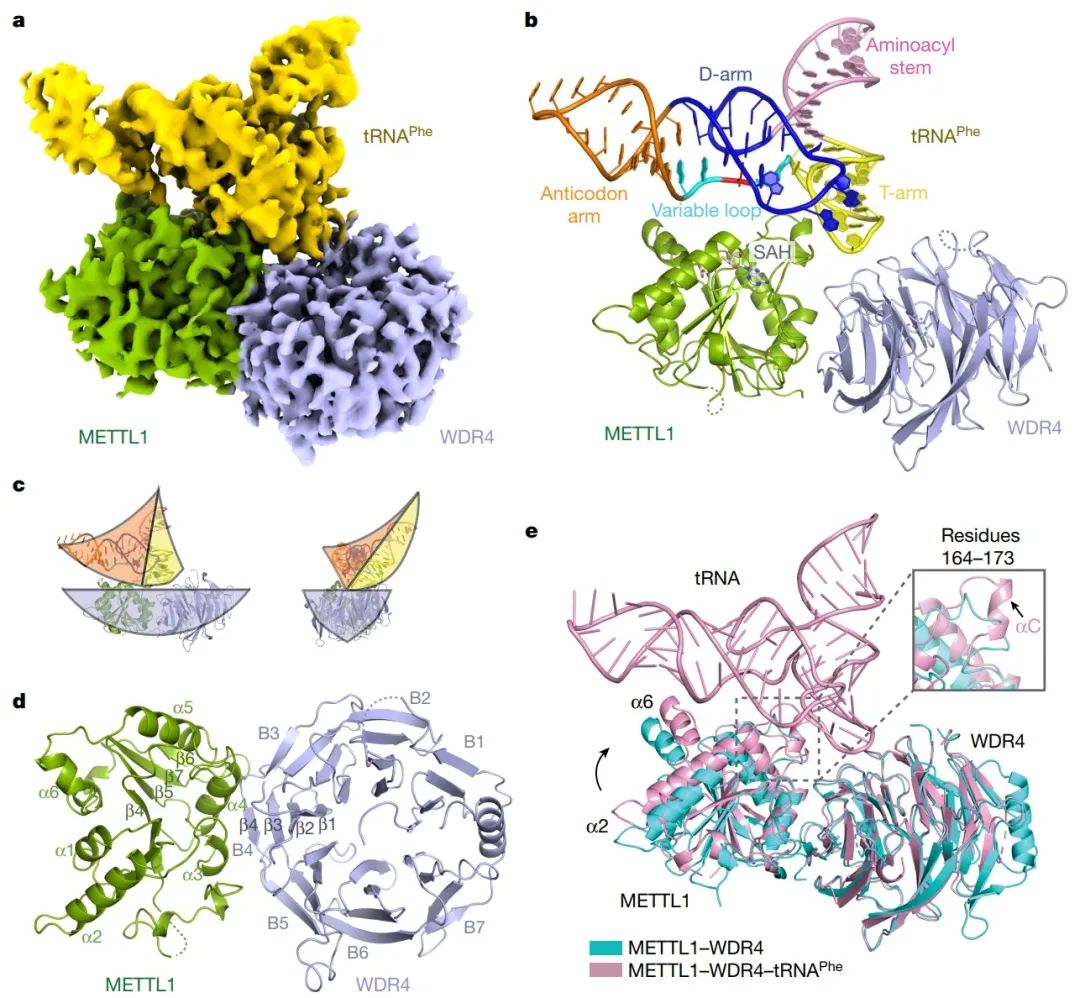
图1. METTL1-WDR4提供特定tRNA附着的平台(图源自Nature )
该研究确定了METTL1 N端(残基1-33)的重要作用,尽管预计它是非结构化的,但在形成MTase催化口袋中起着关键作用。进一步揭示了METTL1 N端结构域的S27磷酸化是如何通过活性位点的空间干扰机制抑制MTase活性的。
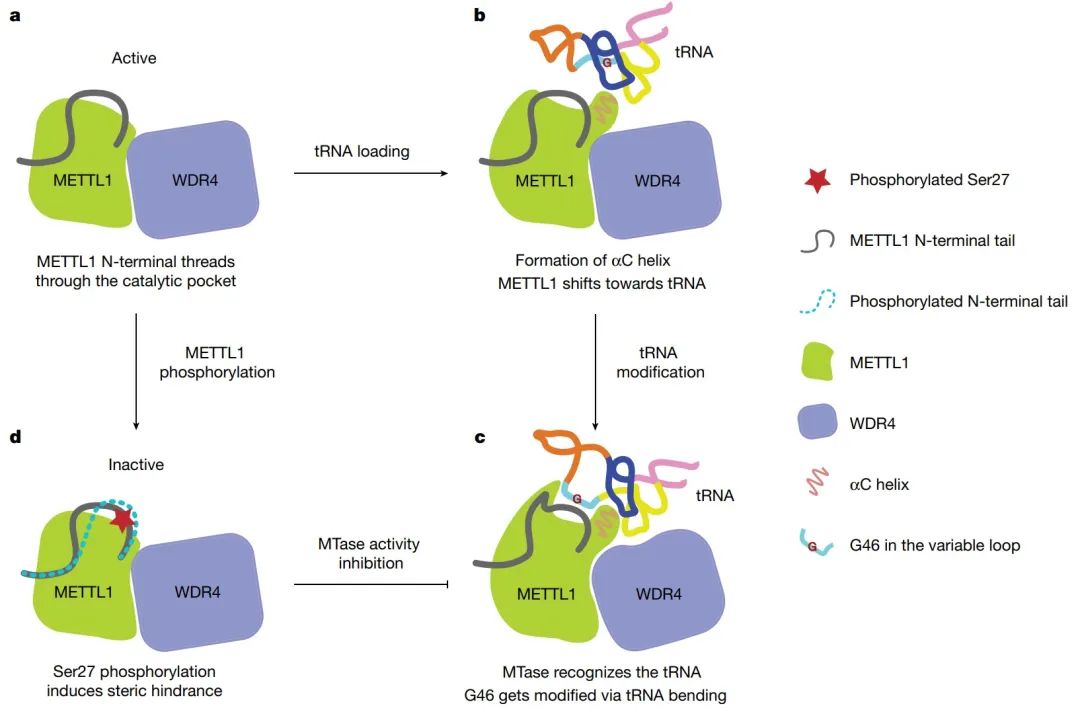
图2. 人METTL1-WDR4底物识别、修饰和催化调控模型(图源自Nature )
总的来说,这项研究揭示了METTL1-WDR4识别tRNA底物的分子机制,强调了METTL1 N端区域在tRNA修饰中的意外作用,并解释了磷酸化介导的MTase活性调控。
原文链接:
https://www.nature.com/articles/s41586-022-05566-4
https://www.nature.com/articles/s41586-022-05565-5

